
 Sahelanthropus tchadensis
Sahelanthropus tchadensisAustralopithecus afarensis
 The first specimens attributed to Australopithecus afarensis were discovered in the 1970’s by Donald Johanson working in the Afar Triangle of Ethiopia at the site of Hadar. The remains from all sites attributed to Au. afarensis have been dated to between 3.9 and 3.0 million years. This is a long time span, indicating that this taxon was a successful species of hominin, persisting for almost a million years. Paleoanthropologists working in Ethiopia and Tanzania over the last 40 years have recovered almost 400 specimens ofAu. afarensis, including fossils from adult and juvenile males and females. These specimens have been invaluable for telling us about the differences between males and females, and for giving insight into how Au. afarensis individuals changed as they aged. Some of this information is coming from a spectacular recent discovery from Dikika in Ethiopia, across the river from Hadar: the relatively complete skeleton of a three year old female. The skull of Au. afarensis is characterized by a prognathic (projecting) face and a relatively small brain. Brain size estimates range from 380-550 cc, which is somewhat larger than the average contemporary chimpanzee. There is a wealth of postcranial bones belonging to this species, including the partial skeleton of Lucy, officially designated AL 288-1. Au. afarensis certainly was bipedal (walked fully upright on two legs), but there has been considerable debate among researchers on exactly how this species walked. The Laetoli footprints, the shape of the pelvis, the curvature of the vertebral column, and the anatomy of the knee all indicate Au. afarensis walked on two legs. However, there are many differences in anatomy between this taxon and modern humans, which have led some paleoanthropologists to argue that Au. afarensis incorporated a degree of arboreal climbing into its locomotor repertoire. and its stride, or gait, was not like ours. These differences include proportionally short legs and longer arms, long, curved finger and toe bones, and subtle differences in the bones of the hip.
The first specimens attributed to Australopithecus afarensis were discovered in the 1970’s by Donald Johanson working in the Afar Triangle of Ethiopia at the site of Hadar. The remains from all sites attributed to Au. afarensis have been dated to between 3.9 and 3.0 million years. This is a long time span, indicating that this taxon was a successful species of hominin, persisting for almost a million years. Paleoanthropologists working in Ethiopia and Tanzania over the last 40 years have recovered almost 400 specimens ofAu. afarensis, including fossils from adult and juvenile males and females. These specimens have been invaluable for telling us about the differences between males and females, and for giving insight into how Au. afarensis individuals changed as they aged. Some of this information is coming from a spectacular recent discovery from Dikika in Ethiopia, across the river from Hadar: the relatively complete skeleton of a three year old female. The skull of Au. afarensis is characterized by a prognathic (projecting) face and a relatively small brain. Brain size estimates range from 380-550 cc, which is somewhat larger than the average contemporary chimpanzee. There is a wealth of postcranial bones belonging to this species, including the partial skeleton of Lucy, officially designated AL 288-1. Au. afarensis certainly was bipedal (walked fully upright on two legs), but there has been considerable debate among researchers on exactly how this species walked. The Laetoli footprints, the shape of the pelvis, the curvature of the vertebral column, and the anatomy of the knee all indicate Au. afarensis walked on two legs. However, there are many differences in anatomy between this taxon and modern humans, which have led some paleoanthropologists to argue that Au. afarensis incorporated a degree of arboreal climbing into its locomotor repertoire. and its stride, or gait, was not like ours. These differences include proportionally short legs and longer arms, long, curved finger and toe bones, and subtle differences in the bones of the hip.Australopithecus sediba
 The most recent addition to the hominin family tree is Australopithecus sediba, named following discoveries made in South Africa at Malapa Cave. “Sediba” is a Lesotho word meaning “fountain” or “wellspring.” Au. sediba is dated to between 1.78 and 1.95 million years ago (ma) using biochronology (a relative dating method utilizjng fossilized non-hominin animals)), paleomagnetism (observing the record of Earth’s magnetic field preserved in magnetic minerals), and uranium-lead (a radioisotoic method that measures the amount of uranium that has decayed to lead in a geologic sample) techniques. Au. sediba has associated cranial (the skull minus the lower jaw) and postcranial (the skeleton minus the skull) remains showing a mosaic of primitive (shared with the ancestral form) and derived (different from the ancestral form) characteristics. This mosaic of features suggests links between Au. sediba and other species in the genus. Australopithecus as well as similarities to species in the genus Homo. Au. sediba is important because it provides insights into hominin variation around the period when the genus Homo emerged. The skull and dentition of Au. sediba display a mosaic of primitive features—in this case, similarities to other species in the genus Australopithecus—and derived features—in this case, features more closely resembling species in the genus Homo. Cranially, the features that link it to Australopithecus include a small cranial capacity (around 420 cubic centimeters), pronounced brow ridges, and enlarged tooth cusps (the pointed eminences on teeth) spaced close together. The discoverers of Au. sediba argue that, among the australopith species, it most closely resembles Australopithecus africanus, which they argue is its likely ancestor. Many differences between Au. sediba and Au. africanus, however, are evident; for example the brain case of Au. sediba is much more vertical on its sides, the temporal lines (which mark the attachment of a chewing muscle that closes the jaw) are widely spaced, its cheek bones are smaller, and its molars are long relative to their width. These features not only set apart this new species from other australopith species, but also link Au. sediba to species in the genus Homo.
The most recent addition to the hominin family tree is Australopithecus sediba, named following discoveries made in South Africa at Malapa Cave. “Sediba” is a Lesotho word meaning “fountain” or “wellspring.” Au. sediba is dated to between 1.78 and 1.95 million years ago (ma) using biochronology (a relative dating method utilizjng fossilized non-hominin animals)), paleomagnetism (observing the record of Earth’s magnetic field preserved in magnetic minerals), and uranium-lead (a radioisotoic method that measures the amount of uranium that has decayed to lead in a geologic sample) techniques. Au. sediba has associated cranial (the skull minus the lower jaw) and postcranial (the skeleton minus the skull) remains showing a mosaic of primitive (shared with the ancestral form) and derived (different from the ancestral form) characteristics. This mosaic of features suggests links between Au. sediba and other species in the genus. Australopithecus as well as similarities to species in the genus Homo. Au. sediba is important because it provides insights into hominin variation around the period when the genus Homo emerged. The skull and dentition of Au. sediba display a mosaic of primitive features—in this case, similarities to other species in the genus Australopithecus—and derived features—in this case, features more closely resembling species in the genus Homo. Cranially, the features that link it to Australopithecus include a small cranial capacity (around 420 cubic centimeters), pronounced brow ridges, and enlarged tooth cusps (the pointed eminences on teeth) spaced close together. The discoverers of Au. sediba argue that, among the australopith species, it most closely resembles Australopithecus africanus, which they argue is its likely ancestor. Many differences between Au. sediba and Au. africanus, however, are evident; for example the brain case of Au. sediba is much more vertical on its sides, the temporal lines (which mark the attachment of a chewing muscle that closes the jaw) are widely spaced, its cheek bones are smaller, and its molars are long relative to their width. These features not only set apart this new species from other australopith species, but also link Au. sediba to species in the genus Homo. Homo rudolfensis
The earliest fossils from the genus Homo are found in eastern, southeastern, and southern Africa. Three species comprise early Homo: Homo rudolfensis (2.5-1.8 million years ago [mya]), H. habilis (2.1-1.5 mya, with which H. rudolfensisshares many similarities) and H. erectus (1.8-0.9 mya). The earliest known species of early Homo, H. rudolfensis fossils are found in Kenya, Ethiopia and northern Malawi. The subject taxon displays an intriguing mix of primitive (traits that are shared with an ancestor) and derived traits (traits different from those found in the ancestral species) that make taxonomic and phylogenetic interpretations difficult and controversial.
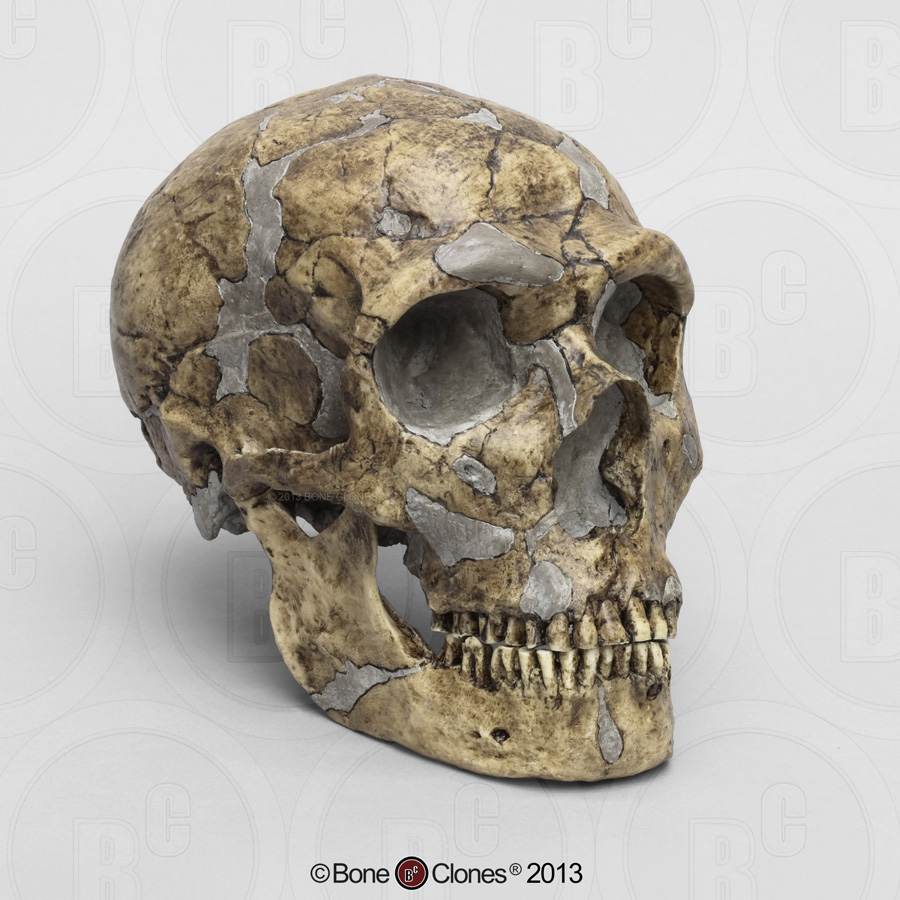 H. rudolfensis is named for the fossil KNM-ER 1470, found on the east side of Lake Rudolf (now Lake Turkana) in Kenya. KNM-ER 1470 most clearly exemplifies H. rudolfensis, and is characterized by a large cranial capacity (around 750 cc), large cheek teeth and a long face that is broad across the orbits (eye sockets) and flattened below the nose. Brain endocasts (a cast made of the mold formed by the impression the brain makes on the inside of the brain case, providing a replica of the exterior surface of the brain) indicate that Broca’s area, an area of the brain associated with speech, is more clearly developed in H. rudolfensis than in australopiths. However, it must be noted that possessing the neurological architecture (i.e., structures within the brain) for speech does not mean that H. rudolfensis used spoken language. In addition to a large brain, H. ridolfensis has very large (i.e., megadont) premolar and molar teeth (i.e., cheek teeth) similar to australopiths, which has led some researchers to question its inclusion in the genus Homo.
H. rudolfensis is named for the fossil KNM-ER 1470, found on the east side of Lake Rudolf (now Lake Turkana) in Kenya. KNM-ER 1470 most clearly exemplifies H. rudolfensis, and is characterized by a large cranial capacity (around 750 cc), large cheek teeth and a long face that is broad across the orbits (eye sockets) and flattened below the nose. Brain endocasts (a cast made of the mold formed by the impression the brain makes on the inside of the brain case, providing a replica of the exterior surface of the brain) indicate that Broca’s area, an area of the brain associated with speech, is more clearly developed in H. rudolfensis than in australopiths. However, it must be noted that possessing the neurological architecture (i.e., structures within the brain) for speech does not mean that H. rudolfensis used spoken language. In addition to a large brain, H. ridolfensis has very large (i.e., megadont) premolar and molar teeth (i.e., cheek teeth) similar to australopiths, which has led some researchers to question its inclusion in the genus Homo.Remains of Homo neanderthalensis have been found at sites throughout Europe, as well as in western Asia. Fossils assigned to this species are also found as far east as Uzbekistan, in Central Asia. The sites from which this speciess is known, which are predominantly cave sites, date from roughly 150 thousand years ago (ka) to as late as roughly 30 ka. Homo neanderthalensis displays many unique features, including features in the skull and postcranial skeleton (skeleton minus skull), which are related to their adaptation to hunting large game in cold environments. Homo neanderthalensisalso had sophisticated stone tool technologies designed to hunt large mammals at close range. This species is important to human evolution because it was contemporary with Homo sapiens and is therefore crucial to our understanding of the origin of our species.
 Many of the unique features possessed by H. neanderthalensis are found in the skull. As with Homo erectus, the general shape of the Homo neanderthalensis skull is long and low with large browridges. Unlike those of H. erectus, however, the browridges of Homo neanderthalensis form individual arches above each eye orbit. (The evolutionary significance of heavy brow ridges, called supra orbital tori, is not certain; scientists are confident they did not act as a visor against the sun, did not contribute to the structural strength of the cranium and were not needed as eye protection. Some researchers suggestb they may have been part of a mate recognition system.) In this way, the Homo neanderthalensis skull resembles that of Homo heidelbergensis. The Homo neanderthalensis skull is quite large, with brain sizes averaging over 1400 cubic centimeters (cc.). Indeed, the brains of Homo neanderthalensis were bigger than those of Homo sapiens; when judged in relation to body size (see below), however, the brain of Homo neanderthalensis is slightly smaller than that of Homo sapiens. The middle and lower parts of the face are positioned far forward relative to the braincase (a condition called “midfacial prognathism”), giving the zygomatics (cheek bones) a “swept back” appearance. The nasal aperture (hole for the nose) in Homo neanderthalensis is very larger, especially when compared to those of Homo sapiens. Because the widest point of the cranium (skill minus lower jaw) is across the middle of the braincase, the skull of Homo neanderthalensis is oval-shaped when viewed from behind (the so-called “en bombe” shape). The braincase also exhibits unique features not found in other hominin species—e.g., occipital buns (thickened, projecting areas at the back of the skull) and suprainiac fossae (small depressions at the back of the skull, just above the occipital bun). The mandibles (lower jaws) are also large and bear molar teeth with large pulp chambers (the area below the enamel in which nerves and blood vessels reside).
Many of the unique features possessed by H. neanderthalensis are found in the skull. As with Homo erectus, the general shape of the Homo neanderthalensis skull is long and low with large browridges. Unlike those of H. erectus, however, the browridges of Homo neanderthalensis form individual arches above each eye orbit. (The evolutionary significance of heavy brow ridges, called supra orbital tori, is not certain; scientists are confident they did not act as a visor against the sun, did not contribute to the structural strength of the cranium and were not needed as eye protection. Some researchers suggestb they may have been part of a mate recognition system.) In this way, the Homo neanderthalensis skull resembles that of Homo heidelbergensis. The Homo neanderthalensis skull is quite large, with brain sizes averaging over 1400 cubic centimeters (cc.). Indeed, the brains of Homo neanderthalensis were bigger than those of Homo sapiens; when judged in relation to body size (see below), however, the brain of Homo neanderthalensis is slightly smaller than that of Homo sapiens. The middle and lower parts of the face are positioned far forward relative to the braincase (a condition called “midfacial prognathism”), giving the zygomatics (cheek bones) a “swept back” appearance. The nasal aperture (hole for the nose) in Homo neanderthalensis is very larger, especially when compared to those of Homo sapiens. Because the widest point of the cranium (skill minus lower jaw) is across the middle of the braincase, the skull of Homo neanderthalensis is oval-shaped when viewed from behind (the so-called “en bombe” shape). The braincase also exhibits unique features not found in other hominin species—e.g., occipital buns (thickened, projecting areas at the back of the skull) and suprainiac fossae (small depressions at the back of the skull, just above the occipital bun). The mandibles (lower jaws) are also large and bear molar teeth with large pulp chambers (the area below the enamel in which nerves and blood vessels reside).The postcranial skeleton of Homo neanderthalensis also exhibits unique features. The entire postcranial skeleton is very heavily-built with thick bones. Individuals were short compared to modern humans; their bodies were also wider, with wider shoulders, rib cages, and hips. The limb bones were short and the distal segments of the limbs (the bones of the forearm and lower leg) were particularly short. These features of the postcranial skeleton are similar to those seen in other mammals that live in cold environments. That is, the skeleton is short and wide to minimize surface area (thereby minimizing heat loss) while maintaining the same mass.
Reference: http://www.becominghuman.org/node/human-lineage-through-time
No comments:
Post a Comment